De genetische codes kunnen bestudeerd
worden vanuit het abstracte standpunt van informatie overdracht bij
het proces van synthese van eiwitten. De codes zijn immers in staat
structuur informatie over te dragen in een omgeving met zeer veel
ruis, storingen, mutaties enz.… Sinds we beschikken over computers
en software en zelf informatie willen communiceren (en daarom moeten
coderen) weten we dat in principe een binaire codering kan gebruikt
worden om gelijk welke willekeurig optredende fout in het
communicatiekanaal te corrigeren. De inzichten die hierbij ontstaan
zijn inspireren verschillende onderzoekers van de genetische code.
Daartoe onderscheiden ze verschillende discrete repertoria (dit
noemen ze verschillende alfabetten) van moleculaire bouwstenen zoals
het 4-letter alfabet van nucleotiden, het 64-letter alfabet van
triplets van die basen, het 20-letter alfabet van de aminozuren die
met deze nucleotiden gesynthetiseerd worden, het 2-letter alfabet van
de sterke en zwakke duplets, en hogere alfabetten van multiplets.
Elke sequentie van nucleotiden kan gelezen worden vanuit een ander
repertorium dat daarmee een ander standpunt mogelijk maakt. Die
kracht herkennen we ook in het haakformalisme: een symbool heeft geen
éénduidige betekenis, een welgevormde haakuitdrukking in twee
symbolen kan indien nodig gelezen worden als haakuitdrukking in
meerdere symbolen. Het moet dus mogelijk zijn om deze onderzoeken in
het haakformalisme te modelleren. We laten ons dus inspireren door
deze onderzoeken, maar geven de onderzoeken een diepere en
fundamentelere basis: het enige dat we nodig hebben is het binaire
onderscheid tussen “iets” en “iets anders”.
Dit blijkt verrassend eenvoudig te zijn wanneer we vertrekken van
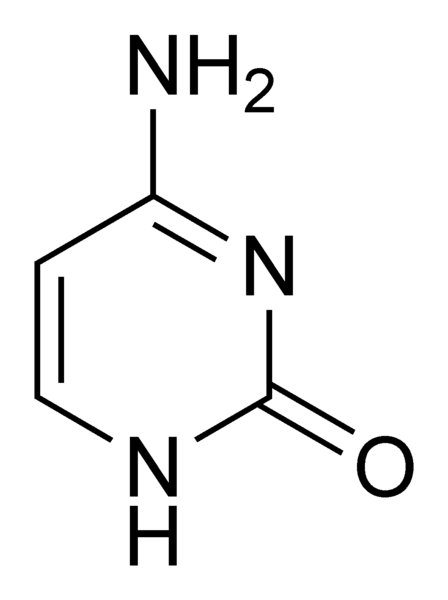
de vier nucleotiden: C, A, G en U/T die de nucleïnezuren RNA en DNA
opbouwen, maar ook een belangrijke rol spelen bij het
energietransport in de cel enz.... U komt voor in het RNA, T komt
voor in het DNA. Het verschil tussen U en T is een -CH3
groep. We herkennen de 3&1 opdeling waarin U/T zich ten opzichte
van de drie andere positioneert. De nucleotide U (en ook zo voor T)
is de enige zonder een -NH2 groep.
In de volgende tabel geven we de namen en een van de isomeren van
de nucleotiden en een deel van de duale classificatie die we nog
verder zullen gebruiken om de structuur in het haakformalisme te
vertalen. Het eerste onderscheid is een tussen een molecule met
enkelvoudige ring (in dit geval een pyrimidine) en een molecule met
een dubbele ring (in dit geval een purine). Het tweede onderscheid is
een classificatie op basis van de reactiviteit van de amino-groep met
HNO2 (reactief, gecodeerd als amino versus niet reactief,
gecodeerd als keto). Beide spelen een rol in de mogelijke mutaties
van α-amino-zuren (zuren met zowel een -NH2 goep als een
-COOH groep op hetzelfde koolstofatoom).
Symbool
|
Naam
|
Pyrimidine
|
Purine
|
Amino
|
Keto
|
Chemische
structuur
|
C
|
Cytosine
|
x
|
|
x
|
|
|
A
|
Adenine
|
|
x
|
x
|
|

|
G
|
Guanine
|
|
x
|
|
x
|

|
U
|
Uracil
|
x
|
|
|
x
|

|
T
|
Thymine
|
x
|
|
|
x
|

|
De vier gematerialiseerde structuren zijn sporen van reacties die
stabiel zijn in het milieu van de cel en als sporen kunnen ze met
elkaar gecombineerd worden waarbij nieuwe sporen ontstaan enz.... Hoe
die allemaal kunnen gecombineerd worden in de opbouw van DNA met zijn
fosfaat-desoxyribose (suiker) ruggengraat en dubbele helix is
voorgesteld in het volgende voorbeeld waarin we links de sequentie
ACGT van boven naar onder lezen en rechts dezelfde sequentie van
onder naar boven:

Het DNA is immers opgebouwd met complementaire strings en de
complementariteit is het gevolg van waterstofbruggen tussen
specifieke baseparen, elektrostatische bindingen tussen molecules die
(gemakkelijker dan de andere bindingen) gemaakt of verbroken worden.
Om de richting van de strings te coderen gebruikt men de standaard
nummering van de ring in de deoxyribose ruggengraat die de
fosfaatgroepen met elkaar verbindt.
Waterstofbruggen zijn enkel te onderscheiden bij duplets.
Waterstofbruggen zijn op het eerste zicht geen binaire
onderscheiding, aangezien een molecule die 3 waterstofbruggen kan
aangaan ook 2 waterstofbruggen kan aangaan, maar omgekeerd is dat
niet mogelijk. Nochtans wordt dat in de natuur niet waargenomen
doordat er ook een ruimtelijke overeenkomst moet zijn tussen de
functionele groepen die waterstof kunnen aanbieden en ontvangen.
Uitzonderingen hierop is het “wobble pair” GU met 2 H-bruggen in
plaats van 3 H-bruggen.
Hierbij de zes mogelijkheden van duplets van nucleotiden, maar
enkel de varianten met een x worden waargenomen:
Waargenomen
|
Duplet
|
Mogelijk aantal waterstofbruggen
|
|
CA
|
2
|
x
|
CG
|
3
|
|
CU
|
2
|
|
AG
|
2
|
x
|
AU
|
2
|
|
GU
|
3
|
Dus enkel de waterstofbruggen tussen CG (namelijk 3) en AU
(namelijk 2) hebben een stabiele structurele functie. Maar er zijn
nog meer ruimtelijke mogelijkheden en onmogelijkheden die dan kunnen
leiden tot nieuwe binaire onderscheiden die een rol kunnen spelen in
de informatieoverdracht van chemische reacties met nucleotiden (een
ervan hebben we al gezien als “wobble pair”). Het zijn dus de
duplets die de driedimensionale ruimte binnenbrengen in de
modellering. Een voorbeeld van de ruimtelijk bepaalde
waterstofbruggen (3 in het linker deel, 2 in het rechterdeel) dank
zij de overeenkomende lengte van de componenten is gegeven in de
schematische voorstelling van een deel van de dubbele string van RNA.

De vier nucleotiden kunnen we dus beschouwen als gebaseerd op drie
binaire onderscheidingen: pyrimidine versus purine, amino versus
keto, en 3 versus 2 mogelijkheden tot waterstofbruggen. Dit brengt
ons tot het volgende overzicht:
Symbool
|
Naam
|
Pyrimidine
|
Purine
|
Keto
|
Amino
|
2
H-bruggen
|
3
H-bruggen
|
C
|
Cytosine
|
x
|
|
|
x
|
|
x
|
A
|
Adenine
|
|
x
|
|
x
|
x
|
|
G
|
Guanine
|
|
x
|
x
|
|
|
x
|
U
|
Uracil
|
x
|
|
x
|
|
x
|
|
T
|
Thymine
|
x
|
|
x
|
|
x
|
|
De vertaling naar onderscheidingen
De binaire onderscheiding pyrimidine versus purine noemen we nu a
versus <a>, keto versus amino noemen we b versus <b>, 2
versus 3 waterstofbruggen noemen we c ten opzichte van <c>. Dus
bijvoorbeeld: cytosine is gemodelleerd als de conjunctie van
pyrimidine, amino, 3H-bruggen of dus als welgevormde haakuitdrukking:
<<a><<b>><<c>>>. Dit is zinvol
omdat we de nucleotiden inderdaad als sporen kunnen afscheiden en ze
elkaar dan uitsluiten, onze meetcontext is zodanig dat we daarin
slagen en dat we er kunnen van uitgaan dat dit betekenis geeft aan
een concatenatie van sporen: een mogelijke genetische code
bijvoorbeeld: ...CUAAGAGUAGG… Die concatenatie wordt dan uitgelezen
door een transfermolecule juist doordat er met behulp van lokaal
stabiele waterstofbruggen twee reactiekernen in contact met elkaar
gebracht worden.
Met de drie binaire onderscheidingen zijn er dus acht mogelijke
toestanden die een reactie kunnen uitlokken in de gepaste context.
Blijkbaar zijn er maar vier gematerialiseerd en als stabiele sporen
beschikbaar binnen het milieu van de cel. We geven een vertaling met
enkel Uracil (we concentreren ons dus nu enkel op de modellering in
mRNA omdat een DNA streng één-op-één afgebeeld wordt om mRNA) die
daaraan voldoet en we voegen een aantal kolommen toe aan de tabel die
relaties verduidelijken.
|
a
|
b
|
c
|
Haakuitdrukking
|
Bitstring
|
Haakvector in een vorm met c als laatst toegevoegde
|
Haakvector met c=<b•a>
|
Haakvector met c=b•a
|
Cytosine
|
0
|
1
|
1
|
<<a>bc>
|
1011.1111
|
a⊕<b>⊕<c>⊕b•a⊕<b•c>⊕c•a ⊕c•b•a
a⊕<b>⊕b•a⊕c•(<>⊕<b>⊕a ⊕b•a)
|
<a>⊕b⊕<b•a>⊕<>
|
<<>>
|
Adenine
|
1
|
1
|
0
|
<ab<c>>
|
1111.0111
|
<a>⊕<b>⊕c⊕<b•a>⊕b•c⊕c•a ⊕c•b•a
<a>⊕<b>⊕<b•a>⊕c•(<<>>⊕b⊕a
⊕b•a)
|
a⊕b⊕b•a⊕<>
|
<<>>
|
Guanine
|
1
|
0
|
1
|
<a<b>c>
|
1101.1111
|
<a>⊕b⊕<c>⊕b•a⊕b•c⊕<c•a> ⊕c•b•a
<a>⊕b⊕b•a⊕c•(<>⊕b⊕<a> ⊕b•a)
|
a⊕<b>⊕<b•a>⊕<>
|
<<>>
|
Uracil
|
0
|
0
|
0
|
abc
|
1000.0000
|
a⊕b⊕c⊕b•a⊕b•c⊕c•a⊕c•b•a
a⊕b⊕b•a⊕c•(<<>>⊕b⊕a⊕b•a)
|
<>
|
<a>⊕<b>⊕<b•a>⊕<<>>
|
Hiermee wordt onmiddellijk duidelijk hoe in het haakformalisme kan
gemodelleerd worden dat maar die vier sporen waargenomen worden: één
van de onderscheidingen is een laatst toegevoegde die dezelfde waarde
heeft als het vectorproduct van de twee andere. De welgevormde
haakuitdrukking is
dus de contractie naar die onderscheiding onder die voorwaarde.
Inderdaad: stel bijvoorbeeld c=b•a (of dus ook a=b•c of b=c•a
of c•b•a=<<>> ) en in de laatste kolommen hebben we
deze gelijkheid en de inbedding gerealiseerd. Dit maakt dan ook
duidelijk dat de realisatie slechts drie sporen als resultaat heeft
in het haakformalisme, want <Uracil> blijkt in die meetcontext
niet anders te zijn dan Adenine (“het ervaren van Uracil is niet
anders dan laten gebeuren van Adenine”). Het vectorproduct is een
universele maar lokale relatie in het haakformalisme, A•C•G•U=<>
betekent dat een 4-vector een van de twee mogelijke waarden heeft,
maar betekent ook A•C•G=<U> dus de gelijkheid wat betreft
waarde van een 3-vector en een 1-vector, of C•G=A•<U>, twee
2-vectoren onderscheiden zich niet.
De nucleotide U blijkt een bijzondere bouwsteen te zijn en dat is
de reden waarom we ervoor kiezen om voor U dezelfde waarde te nemen
voor de drie onderscheidingen. Het is de enige zonder een -NH2
groep en we merken dat Uracil inderdaad door andere kan vervangen
worden om waterstofbruggen te bouwen (bijvoorbeeld door Thymine in
het DNA of Inosine in het tRNA).
Het potentieel universum wordt in die meetcontext slechts door
twee onderscheidingen opgespannen en slechts drie toestanden worden
gerealiseerd zoals in de onderstaande tabel duidelijk wordt. Merk op
dat de vier toestanden elkaar uitsluiten, zowel in drie
onderscheidingen als in twee onderscheidingen en dat ze samen een
tralie opspannen die in het ervaren isomorf is met de
tralie van de laatst toegevoegde onderscheiding.
|
a
|
b
|
c
|
Haakuitdrukking
|
Bitstring
|
Haakvector met c=<b•a>
|
Haakvector met c=b•a
|
Cytosine
|
0
|
1
|
1
|
<<a>bc>
|
1011.1111
|
<a>⊕b⊕<b•a>⊕<>
|
<<>>
|
Adenine
|
1
|
1
|
0
|
<ab<c>>
|
1111.0111
|
a⊕b⊕b•a⊕<>
|
<<>>
|
Guanine
|
1
|
0
|
1
|
<a<b>c>
|
1101.1111
|
a⊕<b>⊕<b•a>⊕<>
|
<<>>
|
<Uracil>
|
1
|
1
|
1
|
<abc>
|
0111.1111
|
<<>>
|
a⊕b⊕b•a⊕<>
|
Dit betekent niet meer en niet minder dat deze vier nucleotiden
een universele uitdrukkingskracht hebben, immers: elke welgevormde
haakuitdrukking kan in twee onderscheidingen uitgedrukt worden en het
gedrag van de nucleotiden wordt door een derde onderscheiding
gemodelleerd. Die derde onderscheiding is een gevolg van de
interactie (en is dus een eigenschap van een duplet, de reactiekern).
Dit maakt onmiddellijk ook de 3&1 structuur duidelijk. Een
uitdrukking zoals bijvoorbeeld C⊕A⊕G⊕<U> is een
welgevormde haakuitdrukking die niet kan onderscheiden worden van het
ervaren van deze relatie. De vier nucleotiden sluiten
elkaar vier-aan-vier uit (het zijn sporen) en dit is duidelijk in
de tabel. De som C⊕A⊕G⊕<U> is in bitstring
1011.1111⊕1111.0111⊕1101.1111⊕0111.1111∼0001.0111. Zo
zijn er 16 welgevormde haakuitdrukkingen wanneer
we elk van deze vier symbolen beschouwen als een 2-vector,
iets wat we altijd kunnen doen. Die 2-vectoren zijn dan in hun meest
eenvoudige vorm zes mogelijke “duplets” die we hieronder gaan
onderzoeken op een abstracter niveau. Inderdaad: als we bijvoorbeeld
een symbool gebruiken als UACCUGAU is de concatenatie van de symbolen
nog niet gemodelleerd. Het enige wat we nu kunnen is individuele
symbolen als sporen in een meetproces realiseren.
Bij realisatie kunnen we drie nucleotiden dus afbeelden op drie
van de vier toestanden in twee onderscheidingen en een van de
nucleotiden op het ervaren supremum. We kunnen kiezen hoe we de
onderscheidingen een naam geven. Een van de onderscheidingen is
gebaseerd op waterstofbruggen en dit is een relatie tussen twee
nucleotiden. Zij vormen samen een duplet. Wanneer we een keten van
nucleotiden beschouwen dan moeten we dit minimaal altijd lezen als
een keten van duplets want twee nucleotiden in dezelfde keten zouden
kunnen reageren met elkaar. Twee identieke nucleotiden vormen ook een
duplet en de volgorde (concatenatie) kan een rol spelen in mogelijke
interpretaties van de string (CA bijvoorbeeld kan onderscheiden
worden van AC, een niet-commutativiteit die we herkennen in het
creatief product). Er zijn dus 42 mogelijke duplets. De 16
duplets kunnen dus 16 welgevormde haakuitdrukkingen modelleren.
Verder zullen we vaststellen hoe essentieel dit is.
In wat volgt moeten we nu een selectie maken van interpretatie van
de nucleotiden in de taal van het haakformalisme en we hanteren de
volgende symbolen met hun voorstelling in drie onderscheidingen:
Vier 1-vectoren
1111.0111∼A
1011.1111∼C
1101.1111∼G
0111.1111∼<U>
Zes 2-vectoren
We construeren alle 2-vectoren en hun combinaties met het
vectorproduct • en dat zowel als haakuitdrukking als in binaire
vorm. We noemen voor het gemak een 2-vector een duplet. Noteer dat
bijvoorbeeld A•<U> niet kan onderscheiden worden van <A•U>
en niet kan onderscheiden worden van <U•A> en dat alle
“duplets” commutatief zijn (een niet commutatieve duplet zou
moeten gemodelleerd worden als een creatief product):
•
|
|
A•C
|
C•G
|
C•<U>
|
A•G
|
A•<U>
|
G•<U>
|
|
•
|
1011.0111
|
1001.1111
|
0011.1111
|
1101.0111
|
0111.0111
|
0101.1111
|
A•C
|
1011.0111
|
1111.1111∼<<>>
|
1101.0111∼A•G
|
0111.0111∼A•<U>
|
1001.1111∼C•G
|
0011.1111∼C•<U>
|
0001.0111∼A•C•G•<U>
|
C•G
|
1001.1111
|
1101.0111∼A•G
|
1111.1111∼<<>>
|
0101.1111∼G•<U>
|
1011.0111∼A•C
|
0001.0111∼A•C•G•<U>
|
0011.1111∼C•<U>
|
C•<U>
|
0011.1111
|
0111.0111∼A•<U>
|
0101.1111∼G•<U>
|
1111.1111∼<<>>
|
0001.0111∼A•C•G•<U>
|
1011.0111∼A•C
|
1001.1111∼C•G
|
A•G
|
1101.0111
|
1001.1111∼C•G
|
1011.0111∼A•C
|
0001.0111∼A•C•G•<U>
|
1111.1111∼<<>>
|
0101.1111∼G•<U>
|
0111.0111∼A•<U>
|
A•<U>
|
0111.0111
|
0011.1111∼C•<U>
|
0001.0111∼A•C•G•<U>
|
1011.0111∼A•C
|
0101.1111∼G•<U>
|
1111.1111∼<<>>
|
1101.0111∼A•G
|
G•<U>
|
0101.1111
|
0001.0111∼A•C•G•<U>
|
0011.1111∼C•<U>
|
1001.1111∼C•G
|
0111.0111∼A•<U>
|
1101.0111∼A•G
|
1111.1111∼<<>>
|
We nemen nu enkel de 2-vectoren uit een van de kolommen of uit een
van de rijen, bijvoorbeeld
1101.0111∼A•G
0111.0111∼A•<U>
1001.1111∼C•G
0011.1111∼C•<U>
Deze vormen (als som in het 3&1 formaat) de welgevormde
haakuitdrukking A•G⊕A•<U>⊕C•G⊕C•U∼0010.1000 of
dus <A•G>. Dat is niet anders dan het creatief product
(<A•G>⊗A•U)A•C∼<A•C<<A•G>>><<A•C><A•U>>∼<A•C<<A•G>>>•<<A•C><A•U>>.
Deze vormen van haakvectoren in 3&1 formaat zijn niet anders dan
uitdrukkingen van disjunctie of conjunctie. Terwijl ook duidelijk is
dat elke 2-vector de disjunctie is van twee nucleotiden
(1101.0111∼A•G is de disjunctie van 1101.1111 (G) en 1111.0111
(A)) omdat de nucleotiden elkaar twee-aan-twee uitsluiten. Die
disjunctie is natuurlijk ook als creatief product (dus in 3&1
formaat) op verschillende manieren uit te drukken, bijvoorbeeld
(<>⊗<A>)G∼<<>>⊕A⊕G⊕<A•G>∼(<>⊗<G>)A∼(<>⊗<A•G>)<A>.
De selectie van 2-vectoren uit een rij (of kolom) levert slechts
vier 2-vectoren op. In het voorbeeld ontbreken A•C en <G•U>
in de selectie, en dat zijn inderdaad de 2-vectoren die we kunnen
beschouwen als toegevoegde voor dezelfde welgevormde haakuitdrukking.
We hebben (<A•G>⊗A•U)A•C al uitgedrukt. Er
geldt bijvoorbeeld ook:
(<A•G>⊗<C•G>)<G•U>∼<<G•U><<A•G>>><<<G•U>><<C•G>>>∼A•G⊕C•G⊕A•<U>⊕C•U∼0010.1000
of dus <A•G>.
Op basis van dezelfde vier 2-vectoren kunnen nog drie ander sommen
gemaakt worden die welgevormde haakuitdrukkingen zijn:
A•G⊕<A•U>⊕<C•G>⊕<C•U>∼1000.1000∼A•U
A•G⊕A•U⊕C•G⊕<C•U>∼0110.0000∼<C•G>
<A•G>⊕<A•U>⊕C•G⊕<C•U>∼1100.0000∼C•U
Elke kolom (rij) heeft een symmetrische kolom (rij) want de
diagonalen geven ofwel <<>>, ofwel <A•C•G•U>,
dus de vormen als sommen in 3&1 formaat geven hetzelfde resultaat
in die symmetrische kolom.
Hiermee is duidelijk dat elke kolom (rij) door middel van een 3&1
som van 2-vectoren elke 2-vector kan representeren. De representatie
is een creatief product en daar zijn dus drie 2-vectoren bij
betrokken. Drie 2-vectoren kunnen als een triplet beschouwd
worden. Een triplet blijkt een fundamentele bouwsteen te zijn in
de genetische code. Een creatief product is niet commutatief en het
is inderdaad bekend dat triplets in mRNA niet commutatief zijn.
We geven nu een keuze van mogelijke triplets waarvan twee
2-vectoren telkens gemeenschappelijk zijn:
(<A•G>⊗A•U)A•C=A•G⊕<A•U>⊕C•G⊕C•U∼0010.1000∼<A•G>
(<A•G>⊗A•U)C•G=A•G⊕<A•U>⊕A•C⊕A•C•G•U∼0000.1000∼<A>
(<A•G>⊗A•U)<C•U>=A•G⊕<A•U>⊕<A•C•G•U>⊕<A•C>∼1010.1000∼A•G•U
(<A•G>⊗A•U)A•G=A•G⊕<A•U>⊕<<>>⊕G•U∼0000.1000∼<A>
(<A•G>⊗A•U)<A•U>=A•G⊕<A•U>⊕<G•U>⊕<>∼1010.1000∼A•G•U
(<A•G>⊗A•U)<G•U>=A•G⊕<A•U>⊕<A•U>⊕<A•G>∼A•U
We zien dat er maar vier verschillende resultaten zijn, en dat er
zowel 1-vectoren, 2-vectoren als 3-vectoren gegenereerd worden. De
triplets die niet gelijk zijn aan een 2-vector worden in dubbel
gegenereerd.
Het aantal mogelijkheden om een geordende keuze te maken van 3
verschillende elementen uit een totaal van 6 elementen, wordt gegeven
door 6!/(6-3)! en dit zijn 120 mogelijkheden waarvan er dus veel
dubbels zullen zijn.
Vier
3-vectoren
Met de vier 1-vectoren A, C, G en <U> kunnen we vier
3-vectoren construeren:
1001.0111∼A•C•G
0011.0111∼A•C•<U>
0101.0111∼A•G•<U>
0001.1111∼C•G•<U>
We merken nu op dat de som in het 3&1 formaat staat en dat:
A•C•G⊕A•C•<U>⊕A•G•<U>⊕C•G•<U>∼1111.1111∼<<>>
De conjunctie van de vier 3-vectoren is ook niet anders dan <<>>,
dus ze sluiten elkaar (enkel als viertal) uit.
De 4-vector
Met de keuze die we maakten voor de vectoren kunnen we nu ook de
4-vector berekenen.
A•C•G•<U>∼0001.0111∼<c•b•a>⊕a⊕b⊕c
Deze welgevormde haakuitdrukking is een zelfduaal op centraal
niveau in drie onderscheidingen en dit is een zeer belangrijke
vaststelling. Zo zijn er 16 zelfduale welgevormde haakuitdrukkingen
in drie onderscheidingen mogelijk:
Bitstring
|
Als
haakuitdrukking
|
Als
som van haakvectoren
|
1010.1010
|
a
|
a
|
0101.0101
|
<a>
|
<a>
|
1100.1100
|
b
|
b
|
0011.0011
|
<b>
|
<b>
|
1111.0000
|
c
|
c
|
0000.1111
|
<c>
|
<c>
|
1001.0110
|
<<c•b•a>>
|
c•b•a
|
1110.1000
|
<<c•b•a>>•<<<<c•a>><<c•b>>>>
|
c•b•a⊕<a>⊕<b>⊕<c>
|
0010.1011
|
<<c•b•a>>•<c•a>c•b
|
c•b•a⊕<a>⊕b⊕c
|
0100.1101
|
<<c•b•a>>•<c•b>c•a
|
c•b•a⊕a⊕<b>⊕c
|
0111.0001
|
<<c•b•a>>•<c•b><c•a>
|
c•b•a⊕a⊕b⊕<c>
|
0110.1001
|
<c•b•a>
|
<c•b•a>
|
0001.0111
|
<c•b•a>•<<<<c•a>><<c•b>>>>
|
<c•b•a>⊕a⊕b⊕c
|
1101.0100
|
<c•b•a>•<c•a>c•b
|
<c•b•a>⊕a⊕<b>⊕<c>
|
1011.0010
|
<c•b•a>•<c•b><<c•a>>
|
<c•b•a>⊕<a>⊕b⊕<c>
|
1000.1110
|
<c•b•a>•<c•b><c•a>
|
<c•b•a>⊕<a>⊕<b>⊕c
|
Maar van die 16 mogelijkheden zijn er maar 10 die een relatie
tussen drie onderscheidingen modelleren. Ze zijn twee-aan-twee
elkaars inbedding. Dus als patroon zijn er maar 5 mogelijkheden. Dus
er zijn 5 mogelijkheden voor de interpretatie van de 4-vector.
A•C•G•<U>∼0001.0111∼<c•b•a>⊕a⊕b⊕c is
maar één mogelijkheid, een andere is <c•b•a>,
een andere c•b•a⊕a⊕<b>⊕c enz…, telkens
overeenkomend met een andere keuze van hoe de onderscheidingen als
symbolen gebruikt worden.
Elke interpretatie genereert dan vier 3-vectoren (die van die
4-vector afgeleid worden) en dus in het totaal zijn er 20 3-vectoren
te onderscheiden en verschillende zullen overlappen (en dus niet
uniek zijn). Dit illustreert wel de robuustheid van de genetische
code en de gelijkwaardigheid van verschillende triplets, maar om het
verband te zien met het bestaan van 20 essentiële aminozuren (die
dus wel uniek zijn) moeten we de structuur nog een stuk verder
ontwikkelen.
De vijf tralies van de 4-vectoren
Met de vier vectoren A, C, G en <U> kunnen we vier
3-vectoren construeren:
A•C•G∼ACG∼1001.0111∼<<c•b•a><<b><a>>>∼<<>>⊕<a>⊕<b>⊕<c>⊕b•a⊕c•a⊕c•b
A•C•<U>∼AC<U>∼0011.0111∼<b<<c><a>>>∼<<>>⊕<a>⊕<c>⊕<b•a>c•a⊕<c•b>⊕c•b•a
A•G•<U>∼AG<U>∼0101.0111∼<a<<c><b>>>∼<<>>⊕<b>⊕<c>⊕<b•a>⊕<c•a>⊕c•b⊕c•b•a
C•G•<U>∼CG<U>∼0001.1111∼<c<<b><a>>>∼<<>>⊕<a>⊕<b>⊕b•a⊕<c•a>⊕<c•b>⊕c•b•a
Dit bevestigt ook dat de som in het 3&1 formaat staat en dat:
A•C•G⊕A•C•<U>⊕A•G•<U>⊕C•G•<U>∼1111.1111∼<<>>
De zes 2-vectoren zijn dan:
A•C∼AC∼1011.0111∼<b<c•a>>∼<>⊕b⊕<c•a>⊕<c•b•a>
A•G∼AG∼1101.0111∼<a<c•b>>∼<>⊕a⊕<c•b>⊕<c•b•a>
A•<U>∼A<U>∼0111.0111∼<ba>∼<>⊕a⊕b⊕b•a
C•G∼CG∼1001.1111∼<c<b•a>>∼<>⊕c⊕<b•a>⊕<c•b•a>
C•<U>∼C<U>∼0011.1111∼<cb>∼<>⊕c⊕b⊕b•c
G•<U>∼G<U>∼0101.1111∼<ca>∼<>⊕a⊕c⊕c•a
Dit maakt duidelijk dat bijvoorbeeld geldt dat
A<<A•C>>∼<<A•C>> maar A•<<A•C>>∼C
A•C•G∼ACG∼1001.0111∼<<A•C>><<A•G>>
Het resultaat is als een volledige tralie af te beelden. We
herkennen de tralie als de structuur van de tralie van het twee
onderscheidingen universum. Het belangrijk verschil is dat geen enkel
punt van de tralie de inbedding is van een ander punt. Elk punt van
de tralie is als creatief product te schrijven en dat creatief
product heeft dus drie componenten.
Niveau
4
|
|
|
|
1111.1111∼<<>>
|
|
|
|
Niveau
3
|
A∼1111.0111∼<ab<c>>∼<a>⊕<b>⊕c⊕<b•a>⊕b•c⊕c•a
⊕c•b•a
|
|
C∼1011.1111∼<<a>bc>∼a⊕<b>⊕<c>⊕b•a⊕<b•c>⊕c•a
⊕c•b•a
|
|
G∼1101.1111∼<a<b>c>∼<a>⊕b⊕<c>⊕b•a⊕b•c⊕<c•a>
⊕c•b•a
|
|
<U>∼0111.1111∼<abc>∼<a>⊕<b>⊕<c>⊕<b•a>⊕<b•c>⊕<c•a>⊕<c•b•a>
|
Niveau
2
|
A•C∼AC∼1011.0111∼<b<c•a>>∼<>⊕b⊕<c•a>⊕<c•b•a>
|
A•G∼AG∼1101.0111∼<a<c•b>>∼<>⊕a⊕<c•b>⊕<c•b•a>
|
A•<U>∼A<U>∼0111.0111∼<ba>∼<>⊕a⊕b⊕b•a
|
|
C•G∼CG∼1001.1111∼<c<b•a>>∼<>⊕c⊕<b•a>⊕<c•b•a>
|
C•<U>∼C<U>∼0011.1111∼<cb>∼<>⊕c⊕b⊕b•c
|
G•<U>∼G<U>∼0101.1111∼<ca>∼<>⊕a⊕c⊕c•a
|
Niveau
1
|
A•C•G∼ACG∼1001.0111∼<<c•b•a><<b><a>>>∼<<>>⊕<a>⊕<b>⊕<c>⊕b•a⊕c•a⊕c•b
|
|
A•C•<U>∼AC<U>∼0011.0111∼<b<<c><a>>>∼<<>>⊕<a>⊕<c>⊕<b•a>c•a⊕<c•b>⊕c•b•a
|
|
A•G•<U>∼AG<U>∼0101.0111∼<a<<c><b>>>∼<<>>⊕<b>⊕<c>⊕<b•a>⊕<c•a>⊕c•b⊕c•b•a
|
|
C•G•<U>∼CG<U>∼0001.1111∼<c<<b><a>>>∼<<>>⊕<a>⊕<b>⊕b•a⊕<c•a>⊕<c•b>⊕c•b•a
|
Niveau
0
|
|
|
|
<A•C•G•U>∼ACG<U>∼0001.0111∼<c•b•a>•<<<<c•a>><<c•b>>>>∼<c•b•a>⊕a⊕b⊕c
|
|
|
|
Zo zijn er dus vijf mogelijke tralies tussen de vijf 4-vectoren en
de gemeenschappelijke <<>>, en de inbedding van die vijf
mogelijke tralies tussen de 4-vectoren en <>.
Overzicht
We construeren nu expliciet de
vijf mogelijkheden. We hebben gekozen om <U> gemeenschappelijk
te nemen voor de vijf mogelijkheden. De ingebedde waarde van U
genereert dan de andere vijf mogelijkheden. Aan de index van de
symbolen kan men de tralie afleiden. Het symbool U geven we dus geen
index. Een gevolg hiervan is dat er maar 7 atomen van de 8 mogelijke
atomen van het drie onderscheidingen universum in de 5 tralies
voorkomen. We komen daar verder op terug.
De tralies zijn ook gemakkelijk te lezen door enkel de bitstrings
te gebruiken. Enkel voor de atomen van de tralies vermelden we in het
overzicht hieronder ook de haakuitdrukking en de haakvector. In drie
onderscheidingen zijn er maar 8 atomen en dus zullen de tralies
overlappen. We geven in de onderstaande tabellen de gelijkheid van
verschillende symbolen aan (gelijkheid die we dank zij de unieke
bitstring kunnen herkennen).
De 2-vectoren en de 3-vectoren geven we enkel weer als
haakuitdrukking in A, C, G, U en als bitstring. Het vectorproduct kan
als inbedding geschreven worden, de disjunctie niet, vandaar dat we
beide noteren.
1
|
a
|
b
|
c
|
Bitstring
|
Haakuitdrukking
|
Haakvector
|
C1
|
0
|
1
|
1
|
1011.1111
|
<<a>bc>
|
a⊕<b>⊕<c>⊕b•a⊕<b•c>⊕c•a ⊕c•b•a
|
A1
|
1
|
1
|
0
|
1111.0111
|
<ab<c>>
|
<a>⊕<b>⊕c⊕<b•a>⊕b•c⊕c•a ⊕c•b•a
|
G1
|
1
|
0
|
1
|
1101.1111
|
<a<b>c>
|
<a>⊕b⊕<c>⊕b•a⊕b•c⊕<c•a> ⊕c•b•a
|
<U>
|
1
|
1
|
1
|
0111.1111
|
<abc>
|
<a>⊕<b>⊕<c>⊕<b•a>⊕<b•c>⊕<c•a>⊕<c•b•a>
|
<A1•C1•G1•U>
|
|
|
|
0001.0111
|
<c•b•a>•<<<<c•a>><<c•b>>>>
|
<c•b•a>⊕a⊕b⊕c
|
De zes 2-vectoren:
A1•C1
|
A1C1
|
1011.0111
|
A1•G1
|
A1G1
|
1101.0111
|
A1•<U>
|
A1<U>
|
0111.0111
|
C1•G1
|
C1G1
|
1001.1111
|
C1•<U>
|
C1<U>
|
0011.1111
|
G1•<U>
|
G1<U>
|
0101.1111
|
De vier 3-vectoren:
A1•C1•G1
|
A1C1G1
|
1001.0111
|
A1•C1•<U>
|
A1C1<U>
|
0011.0111
|
A1•G1•<U>
|
A1G1<U>
|
0101.0111
|
C1•G1•<U>
|
C1G1<U>
|
0001.1111
|
2
|
a
|
b
|
c
|
Bitstring
|
Haakuitdrukking
|
Haakvector
|
C2
|
1
|
0
|
0
|
1111.1101
|
<<c><b>a>
|
<a>⊕b⊕c⊕b•a⊕<b•c>⊕c•a ⊕<c•b•a>
|
A2
|
0
|
1
|
0
|
1111.1011
|
<<c>b<a>>
|
a⊕<b>⊕c⊕b•a⊕b•c⊕<c•a> ⊕<c•b•a>
|
G2
|
0
|
0
|
1
|
1110.1111
|
<c<b><a>>
|
a⊕b⊕<c>⊕<b•a>⊕b•c⊕c•a ⊕<c•b•a>
|
<U>
|
1
|
1
|
1
|
0111.1111
|
<abc>
|
<a>⊕<b>⊕<c>⊕<b•a>⊕<b•c>⊕<c•a>⊕<c•b•a>
|
<A2•C2•G2•U>
|
|
|
|
0110.1001
|
<c•b•a>
|
<c•b•a>
|
De zes 2-vectoren:
A2•C2
|
A2C2
|
1111.1001
|
A2•G2
|
A2G2
|
1110.1011
|
A2•<U>
|
A2<U>
|
0111.1011
|
C2•G2
|
C2G2
|
1110.1101
|
C2•<U>
|
C2<U>
|
0111.1101
|
G2•<U>
|
G2<U>
|
0110.1111
|
De vier 3-vectoren:
A2•C2•G2
|
A2C2G2
|
1110.1001
|
A2•C2•<U>
|
A2C2<U>
|
0111.1001
|
A2•G2•<U>
|
A2G2<U>
|
0110.1011
|
C2•G2•<U>
|
C2G2<U>
|
0110.1101
|
3
|
a
|
b
|
c
|
Bitstring
|
Haakuitdrukking
|
Haakvector
|
C1=C3
|
0
|
1
|
1
|
1011.1111
|
<<a>bc>
|
a⊕<b>⊕<c>⊕b•a⊕<b•c>⊕c•a ⊕c•b•a
|
G2=A3
|
0
|
0
|
1
|
1110.1111
|
<c<b><a>>
|
a⊕b⊕<c>⊕<b•a>⊕b•c⊕c•a ⊕<c•b•a>
|
A2=G3
|
0
|
1
|
0
|
1111.1011
|
<<c>b<a>>
|
a⊕<b>⊕c⊕b•a⊕b•c⊕<c•a> ⊕<c•b•a>
|
<U>
|
1
|
1
|
1
|
0111.1111
|
<abc>
|
<a>⊕<b>⊕<c>⊕<b•a>⊕<b•c>⊕<c•a>⊕<c•b•a>
|
<A3•C3•G3•U>
|
|
|
|
0010.1011
|
<<c•b•a>>•<c•a>c•b
|
c•b•a⊕<a>⊕b⊕c
|
De zes 2-vectoren:
A3•C3
|
A3C3
|
1010.1111
|
A3•G3
|
A3G3
|
1110.1011
|
A3•<U>
|
A3<U>
|
0110.1111
|
C3•G3
|
C3G3
|
1011.1011
|
C3•<U>
|
C3<U>
|
0011.1111
|
G3•<U>
|
G3<U>
|
0111.1011
|
De vier 3-vectoren:
A3•C3•G3
|
A3C3G3
|
1010.1011
|
A3•C3•<U>
|
A3C3<U>
|
0010.1111
|
A3•G3•<U>
|
A3G3<U>
|
0110.1011
|
C3•G3•<U>
|
C3G3<U>
|
0011.1011
|
4
|
a
|
b
|
c
|
Bitstring
|
Haakuitdrukking
|
Haakvector
|
G1=C4
|
0
|
1
|
0
|
1101.1111
|
<c<b>a>
|
<a>⊕b⊕<c>⊕b•a⊕b•c⊕<c•a> ⊕c•b•a
|
G2=A4
|
0
|
0
|
1
|
1110.1111
|
<c<b><a>>
|
a⊕b⊕<c>⊕<b•a>⊕b•c⊕c•a ⊕<c•b•a>
|
C2=G4
|
1
|
0
|
0
|
1111.1101
|
<<c><b>a>
|
<a>⊕b⊕c⊕b•a⊕<b•c>⊕c•a ⊕<c•b•a>
|
<U>
|
1
|
1
|
1
|
0111.1111
|
<abc>
|
<a>⊕<b>⊕<c>⊕<b•a>⊕<b•c>⊕<c•a>⊕<c•b•a>
|
<A4•C4•G4•U>
|
|
|
|
0100.1101
|
<<c•b•a>>•<c•b>c•a
|
c•b•a⊕a⊕<b>⊕c
|
De zes 2-vectoren:
A4•C4
|
A4C4
|
1100.1111
|
A4•G4
|
A4G4
|
1110.1101
|
A4•<U>
|
A4<U>
|
0110.1111
|
C4•G4
|
C4G4
|
1101.1101
|
C4•<U>
|
C4<U>
|
0101.1111
|
G4•<U>
|
G4<U>
|
0111.1101
|
De vier 3-vectoren:
A4•C4•G4
|
A4C4G4
|
1100.1101
|
A4•C4•<U>
|
A4C4<U>
|
0100.1111
|
A4•G4•<U>
|
A4G4<U>
|
0110.1101
|
C4•G4•<U>
|
C4G4<U>
|
0101.1101
|
5
|
a
|
b
|
c
|
Bitstring
|
Haakuitdrukking
|
Haakvector
|
A1=C5
|
0
|
0
|
1
|
1111.0111
|
<<c>ba>
|
<a>⊕<b>⊕c⊕<b•a>⊕b•c⊕c•a ⊕c•b•a
|
A2=A5
|
0
|
1
|
0
|
1111.1011
|
<<c>b<a>>
|
a⊕<b>⊕c⊕b•a⊕b•c⊕<c•a> ⊕<c•b•a>
|
C2=G5
|
1
|
0
|
0
|
1111.1101
|
<<c><b>a>
|
<a>⊕b⊕c⊕b•a⊕<b•c>⊕c•a ⊕<c•b•a>
|
<U>
|
1
|
1
|
1
|
0111.1111
|
<abc>
|
<a>⊕<b>⊕<c>⊕<b•a>⊕<b•c>⊕<c•a>⊕<c•b•a>
|
<A5•C5•G5•U>
|
|
|
|
0111.0001
|
<<c•b•a>>•<c•b><c•a>
|
c•b•a⊕a⊕b⊕<c>
|
De zes 2-vectoren:
A5•C5
|
A5C5
|
1111.0011
|
A5•G5
|
A5G5
|
1111.1001
|
A5•<U>
|
A5<U>
|
0111.1011
|
C5•G5
|
C5G5
|
1111.0101
|
C5•<U>
|
C5<U>
|
0111.0111
|
G5•<U>
|
G5<U>
|
0111.1101
|
De vier 3-vectoren:
A5•C5•G5
|
A5C5G5
|
1111.0001
|
A5•C5•<U>
|
A5C5<U>
|
0111.0011
|
A5•G5•<U>
|
A5G5<U>
|
0111.1001
|
C5•G5•<U>
|
C5G5<U>
|
0111.0101
|
Het uniek laatste atoom
Aan de index van de symbolen kan men de tralie afleiden en sommige
atomen komen in twee tralies voor. Het atoom U hebben we niet
voorgesteld met een index om zijn speciale positie aan te geven. Een
gevolg hiervan is dat er maar 7 atomen van de 8 mogelijke atomen in
de 5 tralies voorkomen.
De onderstaande tabel geeft de mogelijkheid om de 7 een unieke
code te geven (tussen A en G, symbolen niet gerelateerd met de reeds
gebruikte symbolen voor nucleotiden).
1-vector
|
Bitstring
|
7
symbolen
|
<U>
|
0111.1111
|
A
|
<U>
|
0111.1111
|
A
|
<U>
|
0111.1111
|
A
|
<U>
|
0111.1111
|
A
|
<U>
|
0111.1111
|
A
|
C1
|
1011.1111
|
B
|
C1=C3
|
1011.1111
|
B
|
G1
|
1101.1111
|
C
|
G2
|
1110.1111
|
D
|
G2=A3
|
1110.1111
|
D
|
G2=A4
|
1110.1111
|
D
|
A1
|
1111.0111
|
E
|
A1=C5
|
1111.0111
|
E
|
A2
|
1111.1011
|
F
|
A2=A5
|
1111.1011
|
F
|
A2=G3
|
1111.1011
|
F
|
C2
|
1111.1101
|
G
|
C2=G4
|
1111.1101
|
G
|
C2=G5
|
1111.1101
|
G
|
Van de 2-vectoren zijn dat er dan maar 18 van de 28 mogelijke
zoals we in de tabel hieronder weergeven, de laatste tabel geeft de
mogelijkheid om de 18 een unieke code te geven, niet gerelateerd aan
de codes van de vorige tabel:
2-vector
|
Bitstring
|
18
symbolen
|
C1•<U>
|
0011.1111
|
A
|
C3•<U>
|
0011.1111
|
A
|
C4•<U>
|
0101.1111
|
B
|
G1•<U>
|
0101.1111
|
B
|
A3•<U>
|
0110.1111
|
C
|
A4•<U>
|
0110.1111
|
C
|
G2•<U>
|
0110.1111
|
C
|
A1•<U>
|
0111.0111
|
D
|
C5•<U>
|
0111.0111
|
D
|
A2•<U>
|
0111.1011
|
E
|
A5•<U>
|
0111.1011
|
E
|
G3•<U>
|
0111.1011
|
E
|
C2•<U>
|
0111.1101
|
F
|
G4•<U>
|
0111.1101
|
F
|
G5•<U>
|
0111.1101
|
F
|
C1•G1
|
1001.1111
|
G
|
A3•C3
|
1010.1111
|
H
|
A1•C1
|
1011.0111
|
I
|
C3•G3
|
1011.1011
|
J
|
A4•C4
|
1100.1111
|
K
|
A1•G1
|
1101.0111
|
L
|
C4•G4
|
1101.1101
|
M
|
A2•G2
|
1110.1011
|
N
|
A3•G3
|
1110.1011
|
N
|
A4•G4
|
1110.1101
|
O
|
C2•G2
|
1110.1101
|
O
|
A5•C5
|
1111.0011
|
P
|
C5•G5
|
1111.0101
|
Q
|
A2•C2
|
1111.1001
|
R
|
A5•G5
|
1111.1001
|
R
|
Dit wordt ook gereflecteerd in het aantal 3-vectoren die we in de
tabel hieronder weergeven. Er zijn maar zeventien 3-vectoren die zich
onderscheiden van in totaal 56, omdat er vier van hen dubbels zijn en
alle 3-vectoren hebben één bit die dezelfde waarde heeft. De
inbedding van deze zeventien modelleert de 3-vectoren met de
inbedding van Uracil. De laatste tabel geeft de mogelijkheid om de 17
een unieke code te geven, niet gerelateerd aan de codes van de vorige
tabel.
3-vector
|
Bitstring
|
17
symbolen
|
C1•G1•<U>
|
0001.1111
|
A
|
A3•C3•<U>
|
0010.1111
|
B
|
A1•C1•<U>
|
0011.0111
|
C
|
C3•G3•<U>
|
0011.1011
|
D
|
A4•C4•<U>
|
0100.1111
|
E
|
A1•G1•<U>
|
0101.0111
|
F
|
C4•G4•<U>
|
0101.1101
|
G
|
A2•G2•<U>
|
0110.1011
|
H
|
A3•G3•<U>
|
0110.1011
|
H
|
A4•G4•<U>
|
0110.1101
|
I
|
C2•G2•<U>
|
0110.1101
|
I
|
A5•C5•<U>
|
0111.0011
|
J
|
C5•G5•<U>
|
0111.0101
|
K
|
A2•C2•<U>
|
0111.1001
|
L
|
A5•G5•<U>
|
0111.1001
|
L
|
A1•C1•G1
|
1001.0111
|
M
|
A3•C3•G3
|
1010.1011
|
N
|
A4•C4•G4
|
1100.1101
|
O
|
A2•C2•G2
|
1110.1001
|
P
|
A5•C5•G5
|
1111.0001
|
Q
|
Elk van deze zeventien 3-vector atomen op niveau 1 van de tralies
wordt gerealiseerd door drie 1-vector atomen, welke dat zijn is
duidelijk in de naamgeving van de 3-vectoren.
Het unieke gemeenschappelijke atoom kan in deze tralies niet van
waarde veranderen. Het wordt ook niet gemodelleerd wanneer we de vijf
tralies berekenen die gebaseerd zijn op de inbedding van U. Dus dit
atoom kan enkel in een groter universum gemodelleerd worden en daarom
moeten we een onderscheiding bijvoegen bij de reeds gebruikte a, b en
c en dus worden de bitstrings nog eens verdubbeld.
Vier onderscheidingen universum
Om alle mogelijke toestanden te modelleren die sporen kunnen zijn
van processen moeten we dus overgaan naar een universum met 16 atomen
en dus vier onderscheidingen. Het patroon wordt behouden zodanig dat
we nu al weten dat er maar 15 atomen als tralie gemodelleerd zullen
kunnen worden, maar alle atomen in drie onderscheidingen zullen wel
kunnen gemodelleerd worden in vier onderscheidingen. Dit is niet
anders dan het patroon dat we gebruikten toen we met dit onderzoek
gestart zijn (maar dan in drie onderscheidingen): het vectorproduct
van alle gebruikte onderscheidingen heeft een waarde en dat zorgt
voor een contractie naar een universum met één onderscheiding
minder. Dit is de universele manier waarop we in het ervaren een
onderscheidingen universum kunnen contraheren naar een universum met
minder onderscheidingen (en dus ook kunnen expanderen naar een met
meer onderscheidingen). Dat is trouwens ook de manier waarop we
afgeleiden hebben gemodelleerd in het haakformalisme en hiermee
hebben we conjunctie en disjunctie kunnen uitdrukken.
Ook het patroon van dualiteit wordt gewoon verder gezet. Zoals
voor drie onderscheidingen zullen ook in vier onderscheidingen
zelfdualen zich enkel op centraal niveau kunnen bevinden.
Andersdualen bevinden zich daar ook en het hoogste vectorproduct is
nu de andersduaal d•c•b•a. Het patroon van de andere zelfdualen
in drie onderscheidingen, namelijk de vier zelfdualen
c•b•a⊕<a>⊕<b>⊕<c>; c•b•a⊕<a>⊕b⊕c;
c•b•a⊕a⊕<b>⊕c; c•b•a⊕a⊕b⊕<c>, wordt
nu uitgebreid door het vectorproduct met d, maar zowel d, c, b en a
kunnen als “laatst toegevoegde” beschouwd worden. Dus de volgende
uitdrukkingen zijn welgevormd: d•(c•b•a⊕<a>⊕<b>⊕<c>)
maar ook c•(d•b•a⊕<a>⊕<b>⊕<d>) enz…
er zijn dus veel meer relevante 4-vectoren.
Naast de andersduaal d•c•b•a genereert dit ook de volgende
16 andersdualen op centraal niveau:
11101000.00010111∼d•c•b•a⊕<d•a>⊕<d•b>⊕<d•c>
00101011.11010100∼d•c•b•a⊕<d•a>⊕d•b⊕d•c
01001101.10110010∼d•c•b•a⊕d•a⊕<d•b>⊕d•c
01110001.10001110∼d•c•b•a⊕d•a⊕d•b⊕<d•c>
11100001.10000111∼d•c•b•a⊕<c•a>⊕<c•b>⊕<d•c>
00101101.10110100∼d•c•b•a⊕<c•a>⊕c•b⊕d•c
01001011.11010010∼d•c•b•a⊕c•a⊕<c•b>⊕d•c
01111000.00011110∼d•c•b•a⊕c•a⊕c•b⊕<d•c>
11001001.10010011∼d•c•b•a⊕<b•a>⊕<c•b>⊕<d•b>
00111001.10011100∼d•c•b•a⊕<b•a>⊕c•b⊕d•b
01100011.11000110∼d•c•b•a⊕b•a⊕<c•b>⊕d•b
01101100.00110110∼d•c•b•a⊕b•a⊕c•b⊕<d•b>
10101001.10010101∼d•c•b•a⊕<b•a>⊕<c•a>⊕<d•a>
01011001.10100110∼d•c•b•a⊕<b•a>⊕c•a⊕d•a
01100101.10100110∼d•c•b•a⊕b•a⊕<c•a>⊕d•a
01101010.01010110∼d•c•b•a⊕b•a⊕c•a⊕<d•a>
Uiteraard vinden we ook hun inbeddingen terug.
We kunnen dus 16+1=17 infima onderscheiden van tralies met als
supremum <<>>. Elk van deze tralies is af te beelden op
de tralie van een drie onderscheidingen universum.
De uitbreiding naar nog hogere universa ligt dan voor de hand en
we zullen merken dat hetzelfde spoor door meerdere atomen kan
voorgesteld worden. We kunnen dan onmiddellijk afleiden dat de
3-vectoren die als dubbel voorkwamen in drie onderscheidingen, in het
vier onderscheidingen universum als viervoud zullen voorkomen en de
3-vectoren die maar éénmaal voorkomen in drie onderscheidingen, als
dubbel zullen voorkomen. Er zullen ook vectoren uniek zijn voor het
vier onderscheidingen universum.
In de genetische code is inderdaad bekend dat meerdere triplets
kunnen coderen voor hetzelfde aminozuur, het is opvallend dat de 20
essentiële aminozuren ofwel door 4, 2 of 1 triplet gecodeerd worden.
Dit wijst er op dat er inderdaad vier onderscheidingen nodig zijn (en
dus bitstrings van 16 bits) om dan sommige atomen vier maal terug te
vinden.
Wanneer we de 20 essentiële aminozuren zouden willen coderen als
unieke sporen die elkaar uitsluiten, dan is zelfs het vier
onderscheidingen universum niet voldoende, en we moeten overgaan naar
5 onderscheidingen en dus 32 bits. We kunnen dit ook als volgt
benaderen: de dynamiek van vier nucleotiden vereist de modellering in
drie onderscheidingen en de laatst toegevoegde onderscheiding
modelleert wat het spoor zal zijn bij de volgende stap. Dit vereist
in binair formaat 8 bits. Elk van deze bits kunnen we vervangen door
vier bits van het twee-onderscheidingen universum (de som van de vier
nucleotiden in een mogelijk 3&1 formaat). In totaal hebben we dan
32 bits nodig.
Als we dit willen realiseren als sporen die achtergelaten kunnen
worden dan kunnen we niet anders dan overgaan naar triplets. Er zijn
43 mogelijkheden hiervoor, dus 64 en twee maal te veel.
Het blijkt nu dat de 20 aminozuren die essentiëel zijn voor het
leven gecodeerd worden door de 64 mogelijke triplets van nucleotiden,
dat meerdere triplets hetzelfde aminozuur genereren en dat sommige
triplets als start en stop van een proces functioneren. Deze
redundantie zorgt ervoor dat niet alle mutaties van nucleotiden
gevolgen zullen hebben, wat goed is voor de robuustheid van het
systeem voor de gevolgen van willekeurige mutaties.
Zoals we dus relaties gevonden hebben tussen de 4 nucleotiden,
zullen we ook relaties vinden tussen de triplets als we bestuderen
hoe ze gerelateerd zijn met de 20 aminozuren. Dit wordt onmiddellijk
veel complexer omdat het tRNA ook met andere nucleotiden dan deze in
het mRNA koppels kan vormen (het zogenaamde “wobble pairing”),
maar we verwachten dat het patroon zich zal herhalen bij het
toevoegen van nieuwe onderscheidingen bovenop de purine/pyrimidine,
keto/amino en 2H/3H onderscheidingen.
Triplets en individuele nucleotiden worden in het haakformalisme
gemodelleerd door een creatief product en dat is een universele
voorstelling van een welgevormde haakuitdrukking. Een mRNA sequentie
is dan een conjunctie (of disjunctie) die lokaal uitgelezen wordt
door de eenvoudige vergelijking van duplets: een nucleotide van het
mRNA en een van het tRNA.